 打印本文
打印本文  关闭窗口
关闭窗口 水分的平衡调节总是同各种溶质的平衡调节密切联系在一起的,动物与环境之间的水交换经常伴随着溶质的交换。生活在淡水中的鱼不仅要解决水大量渗透到体内的问题,而且还必须不断补充溶质的损失。排泄过程不仅会丢失水分,同时也会丢失溶解在水里的许多溶质。影响动物与环境之间进行水分和溶质交换的环境因素很多,不同的动物也具有不同的调节机制,但各种调节机制都必须使动物能在各种情况下保持体内水分和溶质交换的平衡,否则动物就无法生存。
(一)水生动物的渗透压调节
1.海洋动物 海洋是一种高渗环境,生活在海洋中的动物大致有两种渗透压调节类型。一种类型是动物的血液或体液的渗透浓度与海水的总渗透浓度相等或接近;另一种类型是动物的血液或体液大大低于海水的渗透浓度。
海水的总渗透浓度是1135毫摩尔/千克,与海水渗透浓度基本相同的动物有海胆(Echinus和贻贝(Mytilus)等。这些动物一般不会由于渗透作用而失水或得水,但随着代谢废物的排泄)总会损失一部分水,因此动物必须从以下几个方面摄取少量的水:①从食物中(食物一般含有50~90%的水);②饮用海水并排出海水中的溶质;③食物同化过程中产生的代谢水。由于等渗动物所需要的水量很少,所以一般不需要饮用海水,代谢水的多余部分还要靠渗透作用排出体外。蟹(Maja)等的血液渗透浓度比海水略低一些,这些动物会由于渗透作用失去一些水,它们与等渗动物相比,失水量会稍多一些,但它们也会从食物、代谢水中或直接饮用海水(伴随着排泄溶质)而摄入更多一些的水。还有一些动物的血液或体液的渗透浓度比海水略高一些,如海月水母(Aurelia)、枪乌贼(Loligo)、海蛆(Ligia)、龙虾(Nephrops)、盲鳗(Myxine)、和矛尾鱼(Latimeria)等。对这些动物来说,体外的水会渗透到体内来,渗透速率将决定于体内外的渗透压差。这些动物不仅不需要饮水和从食物和代谢过程中摄取水,而且还需借助于排泄器官把体内过剩的水排出体外。
生活在海洋中的低渗动物(如鲱、鲑、、豹蟾鱼),由于体内的渗透浓度与海水相差很大,因此体内的水将大量向体外渗透,如要保持体内水分平衡,低渗动物必须从食物、代谢过程或通过饮水来摄取大量的水。由于从食物和代谢过程中摄取的水量受到动物对食物需要量的限制,所以饮水就成了弥补大量渗透失水的主要方法。与此同时,动物还必须有发达的排泄器官,以便把饮水中的大量溶质排泄出去(图2-21)。在低渗动物中,排泄钠的组织是多种多样的。硬骨鱼类和甲壳动物体内的盐是通过鳃排泄出去的,而软骨鱼类则是通过直肠腺排出。这些排盐组织的细胞膜上有K+泵和Na+泵,因此可以主动地把钾和钠通过细胞膜排出体外。美洲鳗鲡(Anguilla rostrata)在生活过程中要从淡水迁入海水,尽管外部环境的渗透浓度要发生极大的变化,但它的血液渗透浓度却仍能保持稳定,它对低渗调节的控制是独具特色的。当美洲鳗鲡接触海水时,由于吞食海水并从海水中摄取钠而使血液的渗透浓度增加。接着便出现一些细胞脱水现象,肾上腺皮质增加皮质甾醇(一种激素)的分泌量。这种激素有两个重要作用,一是能使分泌氯化物的细胞从鳃内迁移到鳃的表面,另一是在这些细胞膜内形成大量的Na+泵和K+泵。几天之内钠泵排盐机制便可形成,并能把从海水中摄取的钠排出体外,这样就实现了美洲鳗鲡血液浓度的低渗调节。
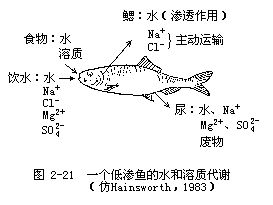
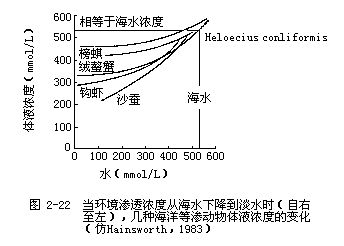
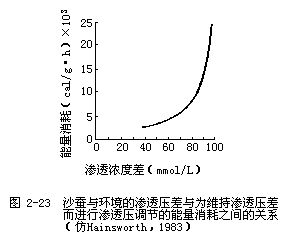
2.低盐环境和淡水环境中的动物 生活在低盐环境和淡水环境中的动物,其渗透压调节是相似的,两种环境只是在含盐量和稳定性方面有所不同。低盐环境(如河海交汇处)的渗透浓度波动性较大,当生活在海洋中的等渗动物游到海岸潮汐区的河流入海口附近时,环境的渗透浓度下降,由于动物与环境之间的渗透浓度差进一步加大,所以动物必须对它们体内的渗透浓度进行调整。图2-22是环境渗透浓度的下降对几种海洋等渗动物体内渗透浓度的影响。当这些动物生活在真正的海水环境中时(图的右上部分),它们的体液浓度都与海水相等或稍高一些。但当环境的渗透浓度下降时(向图中左侧移动),这些动物的体液浓度也不同程度地跟着下降。体液浓度随着环境渗透浓度的改变而改变的动物称为变渗动物;而体液浓度保持恒定,不随环境改变而改变的动物称为恒渗动物。从图2-22可以看到,各种动物调整自身渗透压的精确程度是很不相同的,其中沙蚕(Nereis diversicolor)的体液浓度是下降最多的,当环境浓度是100mmol/L时,沙蚕的体液浓度是200 m mol/L,其浓度差是 100mmol/L。为了维持这样的浓度差,沙蚕用于渗透压调节的能量消耗要增加8倍(图2-23)。但如果靠主动从环境中摄取溶质来维持更大的浓度差,其能量消耗就会如图中曲线所示的那样成指数上升。
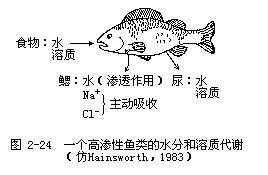
淡水动物所面临的渗透压调节问题是最严重的,因为淡水的渗透浓度极低(约2~3mmol/L)。由于动物血液或体液渗透浓度比较高,所以水不断地渗入动物体内,这些过剩的水必须不断地被排出体外才能保持体内的水分平衡。此外,淡水动物还面临着丢失溶质的问题。有些溶质是随尿排出体外的,另一些则由于扩散作用而丢失。丢失的溶质必须从两个方面得到弥补:一方面从食物中获得某些溶质,另一方面动物的鳃或上皮组织的表面也能主动地把钠吸收到动物体内。钠在数量上是细胞内最重要的一种溶质,其他溶质只依靠从食物中摄取就足够了(图2-24)。
(二)陆生动物的渗透压调节 陆生动物和水生动物一样,细胞内需要保持最适的含水量和溶质浓度。渗透压调节的重要性就在于能保持各种动物细胞内都有相似的含水量,否则细胞的功能就会受影响。
动物失水的主要途径是皮肤蒸发、呼吸失水和排泄失水。丢失的水分主要是从食物、代谢水和直接饮水三个方面得到弥补。但在有些环境中,水是很难得到的,所以单靠饮水远远不能满足动物对水分的需要。因此,陆生动物在进化过程中形成了各种减少或限制失水的适应。陆生动物皮肤的含水量总是比其他组织少,因此可以减缓水穿过皮肤。有很多蜥蜴和蛇,其皮肤中的脂类对限制水的移动发挥着重要作用,如果把这些脂类从皮肤中除去,皮肤的透水性就会急剧增加。
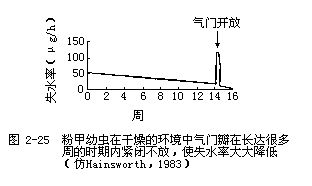
由于水是从动物身体表面蒸发的,所以随着动物身体的减小,其蒸发失水的表面积就会相应增加,这对生活在干燥环境中的小动物,例如陆生昆虫,非常不利。很多陆生昆虫和节肢动物都有特殊适应,尽量减少呼吸失水和体表蒸发失水。例如昆虫利用气管系统来进行呼吸,而气门是由气门瓣来控制的,只有当气门瓣打开的时候,才能与环境进行最大限度的气体和水分交换。如果几个月不喂给粉甲(Tribolium)幼虫食物并把它们置于干燥的空气中,它们的气门瓣常常连续很多星期都紧闭着,气体交换只发生在气门瓣短暂开放的一瞬间(图2-25),这样就可以把蒸发失水量降低到最低限度。节肢动物的体表有一层几丁质的外骨骼,有些种类在外骨骼的表面还有很薄的蜡质层,可以有效地防止水分的蒸发。
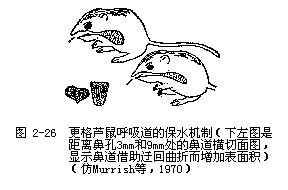
鸟类、哺乳类中减少呼吸失水的途径是将由肺内呼出的水蒸气,在扩大的鼻道内通过冷凝而回收。鼻道温度低于肺表面,来自肺的湿热气遇冷后就会凝结在鼻道内表面并被回收。这种回收冷凝水的工作机制是与许多荒漠鼠类不断吸入干燥的冷空气有关的,当干燥的冷空气通过鼻道时,鼻道表面就会因水分蒸发而变冷,而变冷了的鼻道内表面能使来自肺部的饱含水分的热空气凝结为水,这样就可以最大限度地减少呼吸失水。值得注意的是,世居干燥荒漠的更格芦鼠的鼻道迂回曲折,大大增加了鼻道内的表面积,这是对这一功能的一种形态适应(图2-26)。
减少排泄失水,如许多荒漠鸟兽具有良好重吸收水分的肾脏。人尿中的盐离子浓度比血浆的浓度高3倍,但更格芦鼠尿中的盐浓度却可以比血浆中的高17倍。一般说来,兽类中浓缩尿的能力越强,其肾脏髓质部的相对厚度指数越大,重吸收水的主要部位是位于髓质部中的亨氏袢。许多研究证明,越是栖息于干旱环境的兽类,其肾脏髓质部的相对厚度越大,相应的尿中盐离子浓度比血浆高出的倍数也越大。
含氮废物的排出形式也是减少排泄失水的一种途径。大多数水生生物排出的氮代谢产物是氨(NH4+)。虽然氨也有一定的毒性,但水生生物可以在它达到有害浓度之前就迅速排出体外(主要由鳃排出)。陆生动物则无法为排氮而承受如此大量的水分丧失,因此在蛋白质代谢中常常产出一种毒性较小的代谢产物。哺乳动物所产出的这种氮代谢产物是尿素(CO(NH2)2),由于尿素溶于水,所以排泄过程也会损失一些水分,失水的多少则视肾脏的浓缩能力而定。爬行动物和鸟类则以尿酸(C5H4N4O3)的形式排泄含氮废物,这是对陆地生活的进一步适应。在炎热干燥的沙漠生境中,尿酸甚至可以结晶状态排出体外,这种节水适应可使一些鸟类和爬行动物在沙漠的烈日下也能积极地活动。
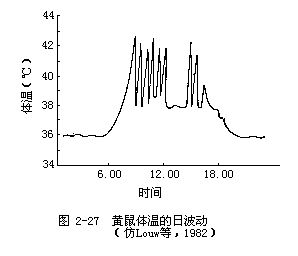
减少呼吸和体表蒸发失水增加了在高温下体温调节的困难,因此,必须靠其他方法加以解决。最普通的一种生理机制是使体温有更大的波动范围(与正常的内稳态动物相比,体温波动幅度要大得多)。例如,黄鼠(Citellus leucurus)体内的酶系统与大多数动物相比,其发挥作用的温度范围要宽得多,因此允许体温有较大幅度的变化。实际上,黄鼠就是靠体温达到极高的水平来解决散热问题的,体温常常比周围环境温度还要高,这样就可维持散热。当体温达到最高点时(42℃),它会躲避到地下洞穴中去降温(图 2-27)。生活在沙漠中的羚羊也有同样的适应,长角羚和瞪羚的体温也常有很大变化。例如,长角羚的直肠温度可达45℃,而瞪羚则可达46.5℃。把身体作为一个热储存器加以利用,可使动物在高温条件下能继续有效地执行各种功能。羚羊的身体比黄鼠更大,因而可以吸收更多的热量,可以长时间地保持活动状态,而不必像黄鼠那样需定期退回洞穴中降温。对羚羊来说,白天所吸收的热量到了较凉爽的夜晚自然就会消散。动物在白天让自己的体温持续不断地升高还有另一种好处,这就是缩小动物体和环境之间的温度差,从而进一步减少动物体的吸热量。对大多数哺乳动物来说,体温超过43℃就会对脑造成损伤。但据观察,瞪羚直肠温度保持46.5℃长达6小时,大脑功能仍完全正常。这是因为血液在到达大脑之前就通过热对流交换使血液降了温,因此羚羊脑的温度比体温要低。
 打印本文 关闭窗口
打印本文 关闭窗口