气体型循环
(一)碳的循环 碳对生物和生态系统的重要性仅次于水,它构成生物体重量(干重)的49%。有机化学就是专门研究碳化合物的一门科学,碳分子独一无二的特性就是可以形成一个长长的碳链,这个碳链为各种复杂的有机分子(蛋白质、磷脂、碳水化合物和核酸等)提供骨架。同构成生物的其他元素一样,碳不仅构成生命物质,而且也构成各种非生命化合物。在碳的循环中我们更加强调非生命化合物的重要性,因为最大量的碳被固结在岩石圈中,其次是在化石燃料(石油和煤等)中,这是地球上两个最大的碳储存库,约占碳总量的99.9%,仅煤和石油中的含碳量就相当于全球生物体含碳量的50倍!在生物学上有积极作用的两个碳库是水圈和大气圈(主要以CO2的形式)(图5-37)。很多元素都与碳相似,有着巨大的不活动的地质储存库(如岩石圈等)和较小的但在生物学上积极活动的大气圈库、水圈库和生物库。物质的化学形式常随所在库而不同。例如,碳在岩石圈中主要以碳酸盐的形式存在,在大气圈中以二氧化碳和一氧化碳的形式存在,在水圈中以多种形式存在,在生物库中则存在着几百种被生物合成的有机物质。这些物质的存在形式受到各种因素的调节。
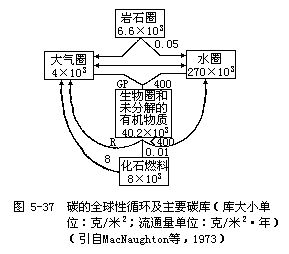
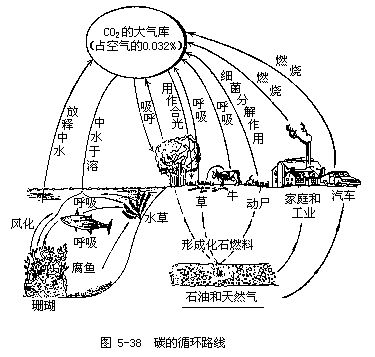
植物通过光合作用从大气中摄取碳的速率和通过呼吸和分解作用而把碳释放给大气的速率大体相等。大气中二氧化碳是含碳的主要气体,也是碳参与循环的主要形式。碳循环的基本路线是从大气储存库到植物和动物,再从动植物通向分解者,最后又回到大气中去。在这个循环路线中,大气圈是碳(以CO2的形式)的储存库,二氧化碳在大气中的平均浓度是0.032%(或320/100万,即320ppm)(图5-38)。由于有很多地理因素和其他因素影响植物的光合作用(摄取二氧化碳的过程)和生物的呼吸(释放二氧化碳的过程),所以大气中二氧化碳的含量有着明显的日变化和季节变化。例如,夜晚由于生物的呼吸作用,可使地面附近大气中二氧化碳的含量上升到0.05%;而白天由于植物在光合作用中大量吸收二氧化碳,可使大气中二氧化碳的含量降到平均浓度0.032%以下。夏季,植物的光合作用强烈,因此从大气中所摄取的二氧化碳超过了在呼吸和分解过程中所释放的二氧化碳;冬季则刚好相反。结果每年4~9月北方大气中二氧化碳的含量最低,冬季和夏季大气中二氧化碳的含量可相差0.002%,即相差20ppm。
除了大气以外,碳的另一个储存库是海洋。实际上海洋是一个更重要的储存库,它的含碳量是大气含碳量的50倍。更重要的是,海洋对于调节大气中的含碳量起着非常重要的作用。在植物光合作用中被固定的碳,主要是通过生物的呼吸(包括植物、动物和微生物)以二氧化碳的形式又回到了大气。除此之外,非生物的燃烧过程也使大气中二氧化碳的含量增加,如人类燃烧木材、煤炭以及森林和建筑物的偶然失火等。正如前面已提到过的,地球上最大的碳储存库是岩石圈,其中包括由生物遗体所形成的泥炭、煤和石油以及由软体动物的贝壳和原生动物的骨骼所形成的石灰岩(主要成分是碳酸钙)。此外,有很多生长在碱性水域中的水生植物,在进行光合作用时会释放出碳酸钙(光合作用的副产品)。例如,伊乐藻(Elodea canadensis)在自然光照条件下每10小时就可释放出相当自身重量2%的碳酸钙。这种纯碳酸钙和粘土混合就可形成泥灰岩,泥灰岩长期受压就可转变为石灰岩。广泛分布于世界各地的石灰岩大都是这样生成的。岩石圈中的碳也可以重返大气圈和水圈,主要是借助于岩石的风化和溶解、化石燃料的燃烧和火山爆发等。
二氧化碳在大气圈和水圈之间的界面上通过扩散作用而互相交换着,而二氧化碳的移动方向决定于它在界面两侧的相对浓度,它总是从浓度高的一侧向浓度低的一侧扩散。借助于降水过程,二氧化碳也能进入水圈。例如,一升雨水中大约含有0.3毫升的二氧化碳。在土壤和水域生态系统中,溶解的二氧化碳可以和水结合形成碳酸(H2CO3),而且这个反应是可逆的。碳酸在这个可逆反应中可以生成氢离子和碳酸氢根离子
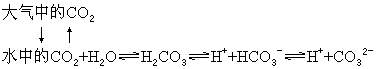
由于所有这些反应都是可逆的,所以反应进行的方向就取决于参加反应的各成分的浓度。由此可以想到,如果大气中的二氧化碳发生局部短缺,就会引起一系列的补偿反应,水圈里的溶解态二氧化碳就会更多地进入大气圈。同样,如果水圈里的碳酸氢根离子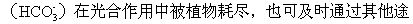 径或从大气中得到补充。总之,碳在生态系统中的含量过高或过低,都能通过碳循环的自我调节机制而得到调整,并恢复到原有的平衡状态。放射性碳(14C)可用来估计空气和水之间二氧化碳的交换速度。由于核武器试验使大气中含有很多的碳同位素。观察空气中14C的减少情况就能计算出二氧化碳在溶于海水以前在大气中滞留了多少时间(大约是5~10年)。大气中每年约有1000亿吨的二氧化碳进入水中,同时水中每年也有相等数量的二氧化碳进入大气。在陆地和大气之间,碳的交换大体上也是平衡的。陆地植物的光合作用每年约从大气中吸收1.5×1010吨碳,植物死后腐败约可释放1.7×1010吨碳。森林是碳的主要吸收者,每年约可吸收3.6×109吨碳,相当其他类型植被吸收碳量的两倍。森林也是生物碳库的主要储存库,约储存着482×109吨碳,这相当于目前地球大气含碳量的2/3。
径或从大气中得到补充。总之,碳在生态系统中的含量过高或过低,都能通过碳循环的自我调节机制而得到调整,并恢复到原有的平衡状态。放射性碳(14C)可用来估计空气和水之间二氧化碳的交换速度。由于核武器试验使大气中含有很多的碳同位素。观察空气中14C的减少情况就能计算出二氧化碳在溶于海水以前在大气中滞留了多少时间(大约是5~10年)。大气中每年约有1000亿吨的二氧化碳进入水中,同时水中每年也有相等数量的二氧化碳进入大气。在陆地和大气之间,碳的交换大体上也是平衡的。陆地植物的光合作用每年约从大气中吸收1.5×1010吨碳,植物死后腐败约可释放1.7×1010吨碳。森林是碳的主要吸收者,每年约可吸收3.6×109吨碳,相当其他类型植被吸收碳量的两倍。森林也是生物碳库的主要储存库,约储存着482×109吨碳,这相当于目前地球大气含碳量的2/3。
但是,碳循环的调节机制能在多大程度上忍受人类的干扰,目前还不十分清楚。由于人类每年约向大气中释放2×1010吨的二氧化碳,使陆地、海洋和大气之间二氧化碳交换的平衡受到干扰,结果使大气中二氧化碳的含量每年增加7.5×109吨,这仅是人类释放到大气中二氧化碳的1/3,其余的2/3则被海洋和增加了的陆地植物所吸收。大气中二氧化碳含量的变化引起了人们的关注,大气二氧化碳的含量在人类干扰以前是相当稳定的,但人类生产力的发展水平已达到了可以有意识地影响气候的程度。从长远来看,大气中二氧化碳含量的持续增长将会给地球的生态环境带来什么后果,是当前科学家最关心的问题之一,对此我们将在第6章作更详细的介绍。
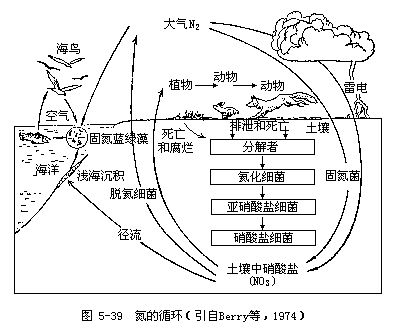
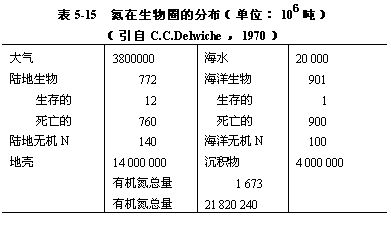
(二)氮的循环 氮是构成生物蛋白质和核酸的主要元素,因此它与碳、氢、氧一样在生物学上具有重要的意义。氮的生物地化循环过程非常复杂,循环性能极为完善(图5-39)。氮的循环与碳的循环大体相似,但也有明显差别。虽然生物所生活的大气圈,其含氮量(79%)比含二氧化碳量(0.03~0.04%)要高得多,但是氮的气体形式(N2)只能被极少数的生物所利用。虽然所有的生物都要以代谢产物的形式排出碳和氮,但几乎从不以N2的形式排放含氮废物。在各种营养物质的循环中,氮的循环实际上是牵连生物最多和最复杂的,这不仅是因为含氮的化合物很多,而且在氮循环的很多环节上都有特定的微生物参加。氮在生物圈内的分布见表5-15。
1.固氮 由于大气成分的79%是氮气,所以氮最重要的储存库就是大气圈,但是大多数生物又不能直接利用氮气,所以以无机氮形式(氨、亚硝酸盐和硝酸盐)和有机氮形式(尿素、蛋白质和核酸等)存在的氮库对生物最为重要。大气中的氮只有被固定为无机氮化合物(主要是硝酸盐和氨)以后,才能被生物所利用。虽然固氮的方法有物理化学法和生物法两种,但其中以生物固氮法最为重要。据估计,靠电化学和光化学固氮,每年平均可固氮7.6×106吨,而生物固氮平均每年的固氮量为54×106吨,人类每年合成氮肥约30×106吨,这也是一个不小的数字。根据人类合成氮肥的增产速度,预计到本世纪末,每年约可生产氮肥100×106吨。C.C.Delwiche认为:现在的工业固氮量约等于现代农业到来之前的生物固氮量。
固氮过程首先需要分子氮激活,使其分裂为两个自由氮原子(N2→2N),这个过程需要消耗能量,在生物固氮时,1摩尔的N2约耗能669×103焦。在自由氮与氢结合形成氨气时(N→NH3),1摩尔的氮气(28克)可释放54×103焦能量,因此,固定1摩尔的氮气,只需输入615×103焦能量就够了(即669-54=615)。除了光化学固氮法外,所有固氮生物都需要从外部提供碳化合物作为一种能源,以便影响这一吸热反应。生物固氮需要两种酶(固氮酶和氢化酶)进行调节,生物固氮的意义在于低能消耗,而工业固氮需要极高的温度和极大的压力(即400℃高温和200个大气压)。
已知有固氮能力的细菌和藻类很多,但为了方便可把它们分为两个类群:一类是共生的固氮生物(主要是细菌,但也有真菌和藻类),另一类是自由生活的固氮生物(包括细菌、藻类和其他一些微生物)。共生的固氮生物主要生活在陆地,而营自由生活的固氮生物在陆地和水域都有。但共生固氮生物在数量上至少要比营自由生活的固氮生物多几百倍。
在共生固氮生物中,根瘤菌(Rhizobium)是最重要的,也是人类了解最清楚的。根瘤菌对宿主植物(如豌豆、三叶草和菜豆等豆科植物)有高度的特异性,一定种类的根瘤菌只同一定种类的豆科植物发生共生关系,这些根瘤菌可潜入豆科植物的根毛,然后进行繁殖。已知有10多种高等植物(如鼠李、杨梅和桤木等)也有共生生物固氮作用。由于豆科植物与根瘤菌之间已经形成了密切的共生关系,所以豆科植物离开了根瘤菌就不能固氮,而把根瘤菌接种在其他植物上也不能固氮。
正如前面已说过的,在水生生态系统中,固氮生物大都是非共生生物,但有一个重要例外,这就是满江红(Azolla)及其共生物蓝绿藻[鱼腥藻(Anabaena)],它们广泛分布于我国温带和亚热带的水稻田中,被农民作为肥料加以利用,因此对农业生产有重要意义。在非共生固氮生物中既有需氧细菌也有厌氧细菌,还包括蓝绿藻。需氧固氮菌[如固氮菌属(Azotobacter)]广泛分布在土壤中以及淡水和海水中;厌氧菌[如梭菌属(Clostridium)]的分布也同样广泛。事实证明:土壤和水体中的很多细菌都有固氮能力,又由于它们数量极多,所以它们固定的氮量也相当可观。例如在盐沼的沉积层中,细菌固氮量相当于藻类固氮量的10倍。这里值得强调的一点是:所有的共生和非共生固氮菌都需要从外部供应糖类,以便作为一种能源去完成固氮过程的吸热反应,因为没有任何一种固氮菌能够进行光合作用(固定1摩尔氮气(N2)需吸热615×103焦)。
为了研究水体中的固氮过程,需把溶于水中的大气氮移出(通常是用氦清除),然后用一种稳定的氮同位素15N取而代之。并且质谱分析仪跟踪观察这种同位素的去向,这种方法与使用14C测定初级生产量的方法大体相同。P.Richard等人用这种方法研究过Sanctuary湖的固氮过程。研究表明:高固氮率与三种蓝绿藻(鱼腥藻属)存在着正相关。对其他两个湖(Mondota和Wingra湖)的研究也表明:高固氮率与其他蓝绿藻(包括Gleotrichia echinulata)的大量存在密切相关。而在马尾藻海,高固氮率则与束毛藻(Trichodesmium属)的存在相关。固氮过程所需要的能量是靠这些蓝绿藻的光合作用提供的,也就是说,蓝绿藻所生产的有机物质提供了固氮所需要的能量(615×103焦/摩尔)。
2.氨化作用 当无机氮经由蛋白质和核酸合成过程而形成有机化合物(主要是胺类,即―NH2)以后,这些含氮的有机化合物通过生物的新陈代谢又会使氮以代谢产物(尿素和尿酸)的形式重返氮的循环圈。土壤和水中的很多异养细菌、放线菌和真菌都能利用这种富含氮的有机化合物。这些简单的含氮有机化合物在上述生物的代谢活动中可转变为无机化合物(氨)并把它释放出来。这个过程就称为氨化作用(ammonification)式矿化作用(mineralization)。实际上,这些微生物是在排泄它们体内过剩的氮。有些具有氨化作用的微生物只能利用胨而不能利用简单的氨基酸,或者只能利用尿素而不能利用尿酸。相反,其他的微生物则能利用多种多样的含氮有机化合物。氨化过程是一个释放能量的过程,或者说是一种放热反应(exothermic reaction)。例如,如果蛋白质的基本构成物是甘氨酸,那么1摩尔的这种蛋白质经过氨化就可释放出736×103焦的热能。这些能量将被细菌用来维持它们的生命过程。
3.硝化作用 虽然有些自养细菌和海洋中的很多异养细菌可以利用氨或铵盐来合成它们自己的原生质,但一般说来,这些含氮化合物难以被直接利用,而必须使它们在硝化作用(nitrifi-cation)中转化为硝酸盐。这个过程在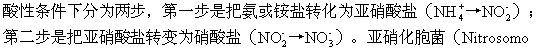
-nas属)可使氨转化为亚硝酸盐,而其他细菌(如硝化细菌)则能把亚硝酸盐转化为硝酸盐。这些细菌全都是具有化能合成作用的自养细菌,它们能从这一氧化过程中获得自己所需要的能量。它们还能利用这些能量使二氧化物或重碳酸盐还原而获得自己所需要的碳,同时产生大量的亚硝酸盐或硝酸盐。据R.Jackson和F.Raw的研究,亚硝化胞菌(N.europaea)每同化一个单位的二氧化碳就可使35个单位的氨氧化为亚硝酸盐;而硝化菌(Nitrobacter agilis)每同化一个单位的二氧化碳可使76~135个单位的亚硝酸盐氧化为硝酸盐。硝酸盐和亚硝酸盐很容易通过淋溶作用从土壤中流失,特别是在酸性条件下。
目前,对开阔海洋及其海底沉积物中的硝化作用还不十分了解。1962年,S.Watson首次报道了从开阔大洋海水中分离出来的海洋亚硝化菌(Nitrosocystis oceanus),他的研究表明,这是一种专性自养细菌,它只能从氨中获得能量和从二氧化碳中获得碳。不少科学家认为,氮素是海洋浮游植物生产量的主要限制因素。
4.反硝化作用(也称脱氮作用) 反硝化作用是指把硝酸盐等较复杂的含氮化合物转化为N2、NO和N2O的过程,这个过程是由细菌[如假单孢菌属(Pseudomonas)]和真菌参与的。这些细菌和真菌在有葡萄糖和磷酸盐存在时可把硝酸盐作为氧源加以利用。大多数有反硝化作用的微生物都只能把硝酸盐还原为亚硝酸盐,但是,另一些微生物却可以把亚硝酸盐还原为氨。在无氧条件下和有葡萄糖存在时,硝酸盐还原为一氧化氮的反硝化过程是一种放热反应,1摩尔的硝酸盐约可放热2.28×106焦。若将1摩尔的硝酸盐还原为分子氮(N2),则可放热2.38×106焦。
由于反硝化作用是在无氧或缺氧条件下进行的,所以这一过程通常是在透气较差的土壤中进行的。依据同样的道理,在氧气含量很丰富的湖泊和海洋表层,反硝化作用便很难发生。但是在水生生态系统缺氧的时期,分子氮就可以通过反硝化过程而产生,这一现象已在Alaskan湖被观察到了。J.Goering从湖底和冰层下1米深处采集水样(冬季),注入标记氮的硝酸盐(K15NO3),然后将水样培养在与湖温相同的温度下。用质谱分析仪进行分析表明:湖底水样中的反硝化过程约比湖面附近快6倍,反硝化作用最重要的终结产物是分子氮,但是没有NO和N2O,分子氮如果未在固氮活动中被重新利用则会返回大气圈库。
5.氮的全球平衡 据估计,全球每年的固氮量为92×106吨(其中生物固氮54,工业固氮30,光化学固氮7.6和火山活动固氮0.2)。但是,借助于反硝化作用,全球的产氮量只有83×106吨(其中陆地43,海洋40和沉积层0.2)。两个过程的差额为9×106吨,这种不平衡主要是由工业固氮量的日益增长所引起的,所固定的这些氮是造成水生生态系统污染的主要因素。最近对海洋环境的研究表明,硝化作用大约可使海洋氮库补充20×106吨氮。从各种来源输入海洋的氮,大体上能被反硝化作用所平衡,基本上能维持一种稳定状态。
至今有一点是很清楚的,即氮的移动绝不是单方向、不可调节和与能量无关的。氮有很多循环路线,而每一条路线都受生物或非生物机制所调节,而且每一个过程都伴随着能量的消耗或释放。氮循环的这许多自我调节机制、反馈机制和对能量的依赖性曾导致提出了这样一个假设,即全球的氮循环是平衡的,固氮过程将被反硝化过程所抵销。目前这一假说还处在讨论之中。如果工业固氮量速率加速增长,而反硝化作用的增加速度又跟不上的话,那么任何已经达到的平衡都可能受到越来越大的压力。另外一个干扰因素是来自汽车和其他机动车所排放的NO2,排放到大气中的含氮气体是造成空气污染的主要原因之一,而且这种污染物对于呼吸系统和大气臭氧层非常有害。在某些生态系统内,硝化过程和反硝化过程能达到极好的协调,以有利于生态系统生产力的需要。这种协调过程在温带地区的冬季进行得最快,以致总是能保证在春季和夏初时使硝酸盐的数量达到最大,而这时也正是植物生长和繁殖需要硝酸盐最多的时候。
|
延伸阅读
|