种的多样性
生物多样性(biological diversity或biodiversity)可定义为“生物中的多样化和变异性以及物种生境的生态复杂性”。它包括植物、动物和微生物的所有种及其组成的群落和生态系统。生物多样性一般有三个水平,即遗传多样性,指地球上生物个体中所包含的遗传信息之总和;物种多样性,指地球上生物有机体的多样化;生态系统多样性,涉及的是生物圈中生物群落、生境与生态过程的多样化。本节仅从群落特征角度来叙述种的多样性,不涉及生物多样性的其他领域。
1.多样性的定义 R.A.Fisher等人(1943)第一次使用种的多样性名词时,他所指的是群落中物种的数目和每一物种的个体数目。后来人们有时也用别的特性来说明种的多样性:比如生物量、现存量、重要值、盖度等。
自从 MacArther(1957)的论文发表后,近几十年来讨论多样性的文章很多,归纳起来,通常种的多样性具有下面二种涵意:
(1)种的数目或丰富度(species richness) 指一个群落或生境中物种数目的多寡。 Poole(1974)认为只有这个指标才是唯一真正客观的多样性指标。在统计种的数目的时候,需要说明多大的面积,以便比较。在多层次的森林群落中必须说明层次和径级,否则是无法比较的。
(2)种的均匀度(species evenness or equitability) 指一个群落或生境中全部物种个体数目的分配状况,它反映的是各物种个体数目分配的均匀程度,例如,甲群落中有100个个体,其中90个属于种A,另外10个属于种B。乙群落中也有100个个体,但种A、B各占一半。那末,甲群落的均匀度就比乙群落低得多。
2.多样性的测定 测定多样性的公式很多,我们这里仅选取其中几种有代表性的作一说明。
(1)丰富度指数 由于群落中物种的总数与样本含量有关,所以这类指数应跟定为可比较的。生态学上用过的丰富度指数很多,现举几例。
a.Gleason(1922)指数:
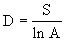
式中A为单位面积,S为群落中物种数目。
b.Margalef(1951, 1957, 1958)指数:
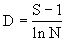
式中S为群落中的总种数,N为观察到的个体总数(随样本大小而增减)。
(2)多样性指数 多样性指数是丰富度和均匀性的综合指标,有人称为异质性指数(Letero-genity indices)或种的不齐性(species heterogenity)。应指出的是,应用多样性指数时,具低丰富度和高均匀度的群落与具高丰富度与低均匀度的群落,可能得到相同的多样性指数。下边是两个最著名的计算公式:
a.辛普森多样性指数(Simpson's diversity index)。辛普森在1949年提出过这样的问题:在无限大小的群落中,随机取样得到同样的两个标本,它们的概率是什么呢?如在加拿大北部寒带森林中,随机采取两株树标本,属同一个种的概率就很高。相反,如在热带雨林随机取样,两株树同一种的概率很低,他从这个想法出发得出多样性指数。所以,
辛普森多样性指数=随机取样的两个个体属于不同种的概率
=1-随机取样的两个个体属于同种的概率
设种i的个体数占群落中总个体数的比例为Pi,那么,随机取种i两个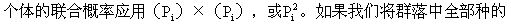 概率合起来,就可得到辛普森指数,即
概率合起来,就可得到辛普森指数,即
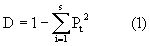
假定我们取样的总体是一个无限总体(在自然群落中,这一假定一般是可以成立的),那么Pi的真值是未知的;它的最大必然估计量是Pi=Ni/N,我们可以用
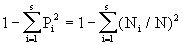
作为总体D值的一个估计量(它是有偏的)。于是
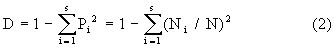
(2)式是实际计算中被采用的公式。
辛普森多样性指数的最低值是0,最高值(1-1/s)。前一种情况出现在全部个体均属于一个种的时候,后一种情况出现在每个个体分别属于不同种的时候。
例如,甲群落中A、B两个种的个体数分别为99和1,而乙群落中A、B两个种的个体数均为50,按辛普森多样性指数计算,则
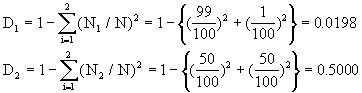
乙群落的多样性高于甲群落。造成这两个群落多样性差异的主要原因是种的不均匀性,从丰富度来看,两个群落是一样的,但均匀度不同。
b.香农-威纳指数(Shannon-Weiner index)。信息论中熵的公式原来是表示信息的紊乱和不确定程度的,我们也可以用来描述种的个体出现的紊乱和不确定性,这就是种的多样性。香农-威纳指数即按此原理设计的,其计算公式为:
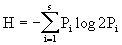
公式中对数的底可取2,e和10,但单位不同,分别为nit,bit和dit,H为信息量(informationcontent),即物种的多样性指数。S为物种数目,Pi为属于种i的个体在全部个体中的比例。
信息量H越大,未确定性也越大,因而多样性也就越高。仍以前面的例子计算,则
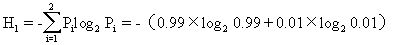
=0.081尼特/个体
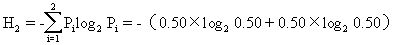
=1.00尼特/个体
可见,乙群落的多样性更高一些,这与用辛普森指数计算结果是一致的。
在香农-威纳多样性指数中包含两个因素:①种类数目,即丰富度;②种类中个体分配上的平均性(equitability)或均匀性(evenness)。种类数目多,可增加多样性;同样,种类之间个体分配的均匀性增加也会使多样性提高。当S个物种每一种恰好只有一个个体时,Pi=1/S,信息量最大,即

均匀度 E=H/Hmax,其中,H实际观察的种类多样性,Hmax为最大的种类多样性。
下面用一个假设的简单数字为例,说明香农-威纳指数的含义。设有A,B,C三个群落,各有两个物种组成,其中各种个体数组成如下:
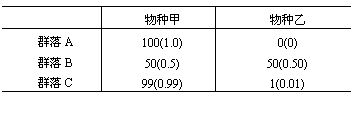
括号内的数字即Pi。因为群落 A的所有个体都属于物种甲,没有任何不定性,从理论上说,H应该等于零,其香农-威纳指数是:
H=-[(1.0×log21.0)+0]=0
由于在群落B中,两个物种各有50个体,其分布是均匀的,它的香农-威纳指数是:
H=-[(0.50×log2 0.50)+(0.50×log2 0.50)]=1
群落 C的两个物种分别具有 99和 1个个体,则:
H=-[(0.99×log2 0.99)+(0.01×log2 0.01)]=0.081
显然,H值的大小与我们的直觉是相符的:群落B的多样性较群落C大(虽然都含两个物种,即丰富程度相等,但群落B异质性程度高),而群落A的多样性等于零。
3.多样性梯度
(1)多样性随纬度的变化 从热带到两极随纬度的增加,物种多样性有逐渐减少的趋势。在乔木、海产瓣鳃类、蚂蚁、蜥蜴和鸟、兽等许多类群中均有充分数据说明这一点,即无论在陆地、海洋和淡水坏境,都有类似趋势。当然也有例外,如企鹅和海豹在极地种类最多,而针叶树和姬蜂在温带物种最丰富。
(2)多样性随海拔的变化 如果在赤道地区登山,随海拔的增高,能见到热带、温带、寒带的环境,同样也能发现物种多样性随海拔增加而逐渐降低。
(3)在海洋或淡水水体物种多样性有随深度增加而降低的趋势 显然,在大型湖泊中,温度低、含氧少、黑暗的深水层,其水生生物种类明显低于浅水区;同样,海洋中植物分布也仅限于光线能透入的光亮区,一般很少超过30米。
我国土地辽阔,南北跨越30余个纬度,由东南向西北干旱度逐渐增加,而西南则是高寒的青藏高原。张荣祖(1985)曾对我国陆生哺乳类(除翼手目外)的种数作过统计比较,发现如下规律:①种数与纬度关系。在北纬40~45°之间,平均种数最低,由40°往更低纬度地区,种数随纬度的降低而增加。②种数与内陆干旱地区的年降水量关系。随着年降水量由50毫米、上升到500毫米,平均种数亦随之而增加。③青藏喜马拉雅-横断山脉地区的种数与海拔高度的关系。随着海拔由850米上升到4750米,平均种数随海拔的升高而降低。
4.决定多样性梯度的因素 为什么热带地区生物群落的物种多样性高于温带和极地?这是由什么因素决定的?对此有不同学说,简介如下:
(1)进化时间学说 热带群落比较古老,进化时间较长,并且在地质年代中环境条件稳定,很少遭受灾害性气候变化(如冰期),所以群落的多样性较高。相反,温带和极地群落从地质年代上讲是比较年轻的,遭受灾难性气候变化较多,所以多样性较低。这就是说,所有群落随时间的推移其种数越来越多,比较年轻的群落可能没有足够的时间发展到高多样化的程度。有些事实能为此学说提供证据,如北半球白垩纪的浮游性有孔虫化石,也和现存有孔虫类一样,从热带到极地,物种多样性逐渐降低。
(2)生态时间学说 考虑更短的时间尺度,认为物种分布区的扩大也需要一定时间。根据这个学说,温带地区的群落与热带的相比是未充分饱和的。从热带扩展到温带不仅需要足够时间,有的种还可能被某种障碍所阻挡,另一些种可能已从热带进入温带。例如牛背鹭就是从非洲经南美而扩展到北美的。
(3)空间异质性学说 当人们由寒带经温带到热带旅行时就能得到一个明显的感觉,环境的复杂性随之而增加。物理环境越复杂,或叫空间异质性程度越高,动植物群落的复杂性也越高,物种多样性也越大。空间异质性有不同的尺度,属于宏观尺度的如地形的变化,山区的物种多样性明显地高于平原区,因为山区有更多样的生境,支持更多样的物种生存。岩石、土壤、植被垂直结构的变化是微观的空间异质性,群落中因这些变化使小生境丰富多样,物种多样性亦高。支持这种学说的证据如群落的垂直结构越复杂,那里的鸟类和昆虫的种类就越丰富。
(4)气候稳定学说 气候越稳定,变化越小,动植物的种类就越丰富,在生物进化的地质年代中,地球上唯有热带的气候可能是最稳定的。所以,通过自然选择,那里出现了大量狭生态位和特化的种类。热带有许多狭食性昆虫,有的甚至只吃一种植物。在高纬度地区,自然选择有利于具广适应性的生物。
(5)竞争学说 在物理环境严酷的地区,例如极地和温带,自然选择主要受物理因素控制,但在气候温和而稳定的热带地区,如热带,生物之间的竞争则成为进化和生态位分化的主要动力。由于生态位分化,热带动植物要求的生境条件往往很狭隘,其食性也较特化,物种之间的生态位重叠也比较多。因此,热带动植物较温带的常有更精细的适应性。
(6)捕食学说 因为热带的捕食者比其他地区多,促使Paine提出捕食说。他认为,捕食者将被食者的种群数量压到较低水平,从而减轻了被食者的种间竞争。竞争的减弱允许有更多的被食者种的共存。较丰富的种数又支持了更多的捕食者种类,Paine认为捕食者促进物种多样性的提高,对于每一营养级都适用。Paine在具岩石底的潮间带去除了顶极捕食动物(海星),使物种多样性由15种降为8种,实验证实了捕食者在维持群落多样性中的作用。
(7)生产力学说 如果其他条件相等,群落的生产力越高,生产的食物越多,通过食物网的能流量越大,物种多样性就越高。这学说从理论上讲是合理的,但现有实际资料有的不支持此学说。例如对丹麦和印度湖泊的枝角类种数与初级生产关系调查结果说明了相反的关系:初级生产力越高,枝角类多样性越低。
热带地区的生长季较长,所以热带群落的种类无论从时间上或空间上分隔环境资源的可能性都较大,从而使共存的种数更多。例如,热带森林鸟类较温带的多,这是因为生产力高的热带森林能提供更多的生存途径;温带森林中没有鹦鹉等食果鸟,没有只吃爬行类的鸟,没有只随蚁群的鸟等等。热带比温带有更丰富的食物来源和营养生态位。
上述7种学说,实际上包括6个因素,即时间、空间、气候、竞争、捕食和生产力。这些因素可能同时影响着群落的物种多样性,并且彼此之间相互作用。各学说之间往往难以截然分开,更可能的是在不同生物群落类型中,各因素及其组合在决定物种多样性中具不同程度的作用。
|
延伸阅读
|