大环境和小环境对生物的不同影响
影响生物生存的非生物因子常常是在相当大的地理区域内起作用的,因此我们可以根据各种物理化学特性划分出不同的地理区域,如根据土壤类型、气候和地质形成过程等。从这种分类中可以得出生态学的一般结论,从而可以知道哪些生物可以在这里定居,哪些生物不能定居。继而,根据生物种类的一定组合特征(即生物群落)可以区分各个不同的气候区,像我们常说的热带森林群落带、温带森林群落带和苔原生物群落带等等。生物群落带(biome)是指具有相似群落的一个区域生态系统类型,它把具有相似非生物环境和相似生态结构的区域连成一个大区。但是这种区分只具有最一般的共性,因为生物只受其邻接环境的影响。例如,森林植物在其下面可提供一个荫凉场所;植物茎和叶的结构和角度可以改变气流并使其下面的地面产生绝热效应。植物的呼吸活动和对气流的阻碍作用都能使湿度和气体浓度发生局部变化。落叶及其所形成的枯枝落叶层在它们腐烂分解以前,会像地毯一样覆盖在土壤表面,起着绝热层的作用。在形成小环境特点方面,动物也起着一定的作用,例如,挖掘地道穴居的动物往往无意中为其他动物创造着可利用的新环境。草食动物的各种取食活动可改变和影响植被的结构,也可创造出小环境。甚至动物排出的粪堆也能影响局部土壤条件,粪堆本身也为食粪动物创造了一个新的小环境。
这些小环境的重要性及其与大环境特征的差异程度可以从Schimitschek的一项研究工作中看出,他研究了一根倒木树干上的小环境分布格局(图2-6),并分析了八齿小蠹(Ips typogra-phus)对树干小环境的利用情况。首先,Schimitschek把树木划分出5个环境特征明显不同的小生境(1~5区),并分析了八齿小蠹在生殖时对这5个生境小区的利用情况。他的结论是,只有第4生境小区能使八齿小蠹进行正常的生殖。第1小区受日光照射太强烈,无法在此产卵;第2小区虽然可以产卵,但受日光照射时间仍然较长,卵会因湿度不足而干瘪;在第3小区,幼虫可以生长发育,但因温度仍然较高而使幼虫在成熟前死亡;第5小区位于倒木的最下面,阴暗潮湿,幼虫死亡率高达75~92%。
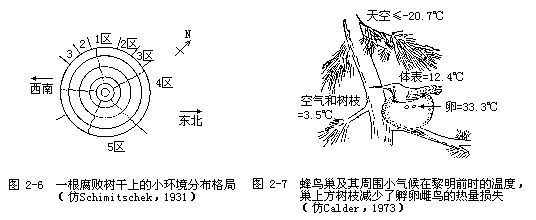
1973年,W.A.Calder研究了小气候与蜂鸟巢的关系,他发现蜂鸟巢的位置总是选择在使卵和雏鸟不致受到不利温度伤害的地方(图2-7)。作为恒温动物的鸟类常因辐射作用而损失体热,蜂鸟巢几乎毫无例外地建筑在一个突出树枝的下方,这个树枝就成了鸟和天空之间的遮护物。此外,鸟巢本身又是一个绝热体,可使鸟卵的温度大大高于孵卵雌鸟身体表面的温度。据估计,如果没有突出树枝的遮护,鸟体辐射损失的热量将会增加大约3倍。如果鸟卵不是放在绝热的鸟巢内和受雌鸟孵卵的话,那么鸟卵的温度到晚上就会接近空气的温度(约4℃左右),而鸟巢内的卵在夜晚时的温度通常都在30℃以上。总之,由于小气候的创造,鸟卵周围环境的温度要比气象所记录的大气候温度高得多。
1969年,M.J.Coe研究了由于巨大的羊茅草(Festuca spp.)草丛的隔离作用而产生的小气候效应。草丛外部空气的温度波动在草丛内部得到了缓冲。据测定:草丛外层叶间的空气温度波动范围是0.3~13.6℃;而在草丛内层叶间的空气温度波动范围是1.8~11.7℃;在羊茅草丛的基部,平均空气温度为7℃,上下波动幅度只有2.1℃。以上研究实例都说明了在生态学工作中,应当特别重视在小环境层次上对非生物因子进行研究。
|
延伸阅读
|