植物的种群生态
一、种群的基本特征
生物很少以孤立的个体形式长期存在,而是以种群形式存在。种群(population)是指一定空间里同种个体的集合,同一种群的个体能自由授粉和繁殖。如某山地的油松(Pinus tabulaeformis)种群,某水域中的水绵(Spirogyra sp.)种群,某农田中的小麦种群等。
种群并不是个体的简单总和,而是一个客观的生态生物学单位,是具有自己独立的特征、结构和机能的整体。一般来说,自然种群具有3个基本特征:① 空间特征,即种群具有一定的分布区域;② 数量特征,即单位面积或体积中的个体数量是动态的;③ 遗传特征,即种群具有一定的基因组成,种群的遗传多样性增加了种群在环境中的生存能力。研究种群数量变动和空间分布规律是种群生态学(population ecology)的主要任务。
种群不仅是物种存在、遗传进化的基本单位,也是群落或生态系统的基本组成单位。
(一)种群的分布
1.种群分布(population distribution)。是指种群在空间中的分布状况,它涉及种群传播、分布类型和格局等要素。此外,种群在特定环境下分布格局的形成,还依赖于种群对其环境的适应性。
物种的分布现状,一方面是其从散布中心或起源中心传播开来的结果,另一方面也是散布的限制因素或生态障碍作用的结果。这些限制因子包括诸如极端温度、积温、湿度等自然气候因子;或像海洋、山脉、陆地等自然地理因子;也有生物因子,有时会因为缺乏某种传粉昆虫而使某种植物无法在这个生境分布。
2.种群分布类型(population distribution type)。是指种群在空间分布的方式。此空间是指一个种群在其所有广大分布范围内的空间,称之为外分布型。种群的分布类型分为连续的(continuous)和间断的(disjunction)或不连续的(discontinuous)的极端类型。
最极端的间断分布是所谓的岛式模型(island model)或岛式分布(island distribution),岛中每一种群各具特色,界限分明,彼此隔离。这不仅是指被水体隔离的岛屿种群,而且也指生长在一系列湖泊、池塘、沼泽包围中的种群,还包括被草原包围的小块森林,或被森林包围的小块草原等生境空间的种群。一个种或种群分布在有利生境空间,而被其它不利生境空间隔离开的任何情况都称为岛式模型。
连续分布是在生境一致的广大空间的种群分布。例如一望无际的大草原、成片的大森林等。实际上这类表面看来似乎是一致的生境也可能不一致,尤其是由于距离上的隔离,连续分布种群也可能无法达到真正的随机交配。
线性分布(linear distribution)是另一种连续分布类型,例如河岸、海岸等生境空间,其种群通常是呈线形连续分布。然而,就整个水系来说则可兼有连续分布和间断分布两重性质,因为通常不同支流上游地区的种群彼此间是空间隔离的。
脚踏石模型(stepping stone model)或脚踏石分布(stepping stone distribution)是含有岛式分布和线性分布的种群分布类型。
显然,大多数种群分布是连续的,造成间断分布通常是由于种群从某地向其它地区远距离散布后,在两地之间缺乏适宜的生境,或者说间隔分布可能代表了连续分布模式的残余,即从前有过宽广的连续分布,但后来由于中间地区消失而造成间断分布。当然,要在这两种极端分布作出区分十分困难。还有一种可能是,种群独立产生于各自不同的分布空间,因而表现出演化上的并行和趋同。同时,某些种群的间断分布,可能不是真正的间断分布,而是代表了人为引种或干预的结果。种群的连续分布和间断分布是一种自然现象,而种群是动态单位,在生理上和遗传上严格地适应其环境,并对环境条件的任何变化都很敏感,而在一定限度内对这些变化发生反应,种群分布类型也是这种生态反应的结果。
3.种群中个体的空间分布格局。是指组成种群的各个体在水平空间中的分布方式或配置状况。分布格局(distribution pattern)是指在一个地理分布区内某一种群各个体的分布状况,也叫种群的内分布型。种群的分布格局,显然是由种群的生物学与生态学特性、种群内及种群间的关系、环境条件的综合影响决定的。在某种意义上它与环境条件的相关是因果关系,或者说种群分布格局是种群对环境长期适应和选择的结果,因而种群分布格局通常反映着一定环境因子对个体行为、生存和生长等的影响。
了解种群的分布格局,对选择估计种群密度的方法有重要意义,也有助于了解种群的动态变化,提供人工栽培和进行种群管理的某些信息。
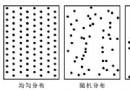
(1)随机分布(random dispersion)。种群个体分布是偶然的,分布机会相等,个体间彼此独立,任一个体的出现与其它个体是否已经存在无关。
出现随机分布的条件:生境条件对很多种的作用差不多;某一主导因素呈随机分布;生境条件比较一致。
(2)均匀分布(uniform dispersion)。种群内的个体分布是等距离,或个体间保持一定的均匀间距,取样时空白和密度大的样方都少,接近平均株数的样方多。
均匀分布在自然条件下极为少见,大多为人工群落,并且以植物种群为多见。人工栽培的株行距一定的种群是典型的均匀分布。
引起均匀分布的原因有:种内竞争;优势种成均匀分布而使其伴生植物也成均匀分布;地形或土壤物理性状的均匀分布;虫害引起;自毒现象。
(3)聚群分布(aggregated dispersion)。种群个体的分布极不均匀,常成群、成簇、成块、斑点状密集分布,各群的大小、群间距离、群内个体的密度等都不相同,但各群大多呈随机分布,有时各群间亦均匀分布。
聚群分布是最广泛的一种分布格局。形成聚群分布的原因有:种的繁殖特点,如植物从母株上散布种子,落在附近,种子长成植株,形成群状;有些植物果实内含有多粒种子,如松树球果,这些种子长成植株总是非常靠近,呈簇状;植物的无性繁殖,形成密集的团聚,如伐根萌芽和根蘖形成的集群。环境中局部条件的差异,如林地的光照条件呈大小不等的光斑镶嵌,郁闭林内的森林更新,多发生在光斑处;局部地形的微起伏和土壤条件的局部差异。种间相互关系,如种间信息等,有可能是直接的有利作用或间接的互为环境。聚群分布能更好地改善小气候与微气候。
种群的分布类型如图13-3所示。有时在同一群落内的某一种群,可以形成两种分布格局。如某一种群侵入某一生境,种子自然撒播可能形成随机分布,随后由于无性繁殖形成聚群分布,最后又因竞争或其它原因呈随机分布或均匀分布。有时候同一种群与不同种生长在一起时,可以形成不同的分布格局。有时候同一种群在不同分布区内保持相同的分布格局。
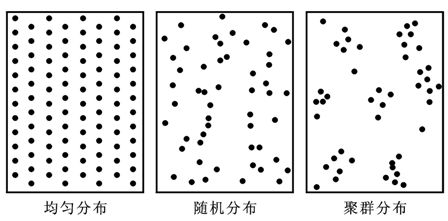
图13-3 种群的三种分布类型
(二)种群的数量及增长
种群数量是指在一定面积或容积中某一个种的个体总数。一个种群的个体数目多少,也叫种群大小(population size)。如果用单位面积或单位容积内的个体数目来表示种群大小,则称为种群的密度(population density)。种群的数量决定于出生率和死亡率的对比关系。单位时间内出生率与死亡率之差为增长率(growth rate)。设种群的起始数量为N0,单位时间后种群的数量增加到N1,则有限增长率(周期增长率)(finite rate of increase):
λ= N1/N0
1.指数式增长。如果种群的个体之间没有竞争,环境资源是无限的,种群数量将呈指数式增长,增长曲线为J型(图13-4)。图中曲线可以表示为:
Nt+1=λNt 或Nt+1= N0λr 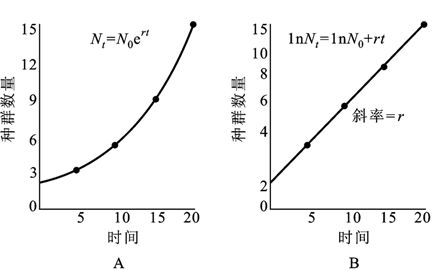
图13-4 种群的指数式增长
A.算术标尺 B.对数标尺
如果所研究的生物在每年的生长季节都连续不断地繁殖,并且种群是世代重叠的,其种群增长可以用Lotka(1922)提出的方程式表示:
dN/dt=rN
其积分式为:Nt = N0ert
其中r称为种群的瞬时增长率(instantaneous growth rate),它与周期增长率λ的关系为:
r=lnλ 或λ=er
参数r值随不同种群的大小、状况及其所处的环境而变化。为了比较不同种群的r值,常把理想状态下的种群能到达的最大增长率称为内禀增长力(innate capacity of increase),用rm表示。rm也被称为生物潜能(biotic potential),其价值在于作为对野外种群研究的一个标准,以比较野外种群与其最大理论增长率之间的差异程度,而这种差异则被看作是由于“环境阻力”造成的。
2.逻辑斯蒂增长。环境资源总是有限的,所以种群不可能长期连续呈指数式增长。随着种群个体数量增加,对于有限空间和其它生活必需资源的种内竞争也加剧,这必然影响到种群的出生率和存活率,从而降低种群的实际增长率。当种群个体的数目接近于环境所能支持的最大值,即环境负荷量(carrying capacity)K值时,种群将不再增长而保持在该值左右,这时dN/dt=0,Pearl and Reed (1920)提出了描述这种种群增长过程的方程式,即逻辑斯蒂方程(Logistic equation):
dN/dt =r N(K-N)/K
逻辑斯蒂曲线图形如图13-5所示,呈一条向着环境负荷量(K)逼近的“S”形增长曲线。它反映了自然种群增长的普遍规律,但对于寿命长的、世代重叠多的种群会产生很大偏差,只有低级生物,如细菌、酵母或浮游动植物才与之比较吻合。此处,(K-N)/K表示环境阻力。当K-N>0时,种群增长;K-N<0时,种群个体数目减少;当K-N=0时,种群数量保持稳定状态。
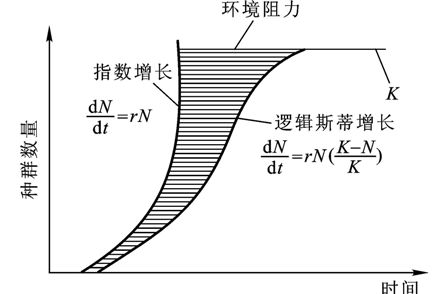
图13-5种群逻辑斯蒂增长的理论曲线
(三)种群密度
种群密度(population density)是指单位面积上的个体数目或种群生物量。密度的计数个体为个体/基株、构件、宿主等。根据计数空间不同,生态学上常区分粗密度和生态密度。粗密度(crude density)是指单位总空间的个体数,即单位面积或容积按种的分布区计算;而生态密度(ecological density)是指在单位栖息空间(种群实际上占据的有用面积或空间)的个体数。在进行密度调查与对比分析时,还需要区分出绝对密度和相对密度。绝对密度是指单位面积或单位空间内种群大小的绝对数量。而相对密度是指单位面积或单位空间内种群大小的相对数量,有时可用时间表示,也可用各种百分比表示。
种群密度的计测方法:
(1)总数量调查法:计数某地段中某种生物个体的全部存活者数量,然后将其总个体数除以该地段总面积,即为其绝对密度。该方法多用于大型的、易被发现的或集中群居的生物。
(2)取样调查法:一般植物调查多用取样方法。具体方法是:根据种群内个体的分布格局确定抽样方法,选择一定数量的样方,在选定样方中计数某种群的全部个体,然后用统计方法求出平均数,将其平均数推广用以估计种群的整体数量。
(四)年龄结构
种群年龄结构是指种群内个体的年龄分布状况。即不同年龄的个体在种群内的比例或配置情况。若按年龄由小到大将其比例绘制成图,其形状类似于金字塔,故又称为年龄金字塔或年龄锥体。
归纳种群的年龄结构,可把种群分为增长型、稳定型和衰退型等三种类型(图13-6)。
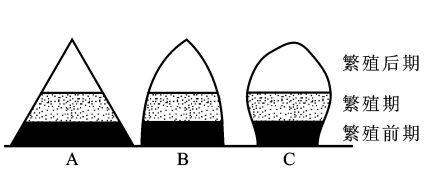
图13-6 种群年龄结构的类型 (Komondy,l976)
A增长型 B稳定型 C衰退型
1.增长型种群(increasing population)。增长型种群以幼年个体占有最大比例,老年个体数最少。幼、中年个体除了补充死去的老年个体外还有剩余,所以,这类种群的数量呈上升趋势。
2.稳定型种群(stable population)。稳定型种群各个年龄级的个体数分布比较均匀,每个年龄级进入上一级的个体数,与下一个年龄级进入该级的个体数大致相等,所以,种群大小趋于稳定。
3.衰退型种群(declining population)。衰退型种群与增长型种群相反,老年个体数大,幼年个体数小,大多数个体已过了生殖年龄,种群的数量趋于减少。
值得注意的是,在利用种群年龄结构分析其发展趋势时,还应就环境因子对种群的作用加以考虑,因为有时幼苗的数量并不能完全说明种群的发展趋势,如在亚热带松-栎混交林中,松的幼苗常比栎多,但由于栎的耐阴性比松强,所以,在郁闭林下,不利于喜阳的松苗生长,而有利于栎苗生长,尽管松的幼苗多,但最终被栎所替代。
种群的年龄结构不仅反映了种群不同年龄个体的组配情况,也反映了种群数量动态及其发展趋势,并在很大程度上反映了种群与环境间的相互关系以及它们在群落中的作用和地位。结合种群的生态需求、各龄级的死亡率和产生后代的能力,就能更好地对种群未来作出估计。
(五)种群大暴发或大发生与崩溃
具不规则或周期性波动的生物都可能出现种群大发生。具该种形式的种群常开始呈指数式增长,后由于各种原因往往出现大批死亡而使种群崩溃(crash)。
浮游植物的赤潮与开花是一个典型例子。赤潮是指水域中一些浮游生物(如腰鞭毛虫、裸甲藻、梭角藻、夜光藻等)暴发性增殖引起水色异常的现象,主要发生在近海,又称红潮。其主要成因是有机污染,氨、磷等营养物过多形成富营养化。
水生植物暴发造成危害的例子也不鲜见。槐叶萍(Salvinia molesta)原产巴西,1952年首次在澳大利亚出现,由于它每2.5d就能加倍,迅速增殖并扩散。到1978年覆盖了昆士兰一个湖泊的400公顷面积,总重达50000t,造成对交通、灌溉和渔业的严重危害。直到1980年由巴西引进专食槐叶萍的象鼻虫(Cyrtobagous sp.)才将危害控制住。具种群暴发现象的水生植物还有伊乐藻(Elodea canadensis)和凤眼莲(Eichornia crassipes)。虽然在国外通常把这些水生植物大发生作为危害,但我国农村还常将凤眼莲等用作家畜饲料进行定期收获。此外,水葱(Scirpus validus)、凤眼莲、绿萍(Lemna minor)、金鱼藻、菱角(Trapa bicornis)等有吸收水中重金属元素的作用,有的还能对水中悬浮物、氯化物、有机氮等有一定净化能力。
陆生植物如贯叶金丝桃(Hypericum perforatum)是一种有毒的多年生杂草,广布于欧亚大陆。在过度放牧的草场上,牛羊喜食的草本植物减少,贯叶金丝桃成了重要危害。当牲畜取食少量时,刺激口舌,降低食欲,大剂量则毒害致死。据报道,1904年被带入美国加州北部,到1944年,扩展到80万公顷面积。用化学药物虽能杀死它,但耗资巨大。直到引进双金叶甲(Chrysolina quadrigemina)才得以控制。此叶甲的幼虫在冬季啃食叶基,使其次春不能长叶,于是根不能贮存营养物,3年后在干旱夏季中死亡。
(六)种群的遗传与进化
植物所具有的表现型特征从根本上说都是由植物内在基因型控制的,同时也受环境条件影响。任何一个种群,其各个个体基因型的相对稳定性是种群繁殖的基础。但是,各个个体的基因型并不完全相同,因而它们各自的表现型在很多情况下也出现差异。种群内在的生存和繁殖差异,使得那些能较好适应环境的个体产生更多后代,结果使种群更适应环境。如果环境条件随时间发生变化,优良的基因型将能较好地适应新环境,并在自然选择中种群的遗传组成将发生变异,从而产生适应性更强的表现型。物种或种群的遗传组成随时间而发生变化的过程就是进化(evolution)。
大多数植物并不都生长在环境条件完全相同的地方,而是有一个环境条件差异较大的地理分布范围。由于长期受到不同环境条件的影响,同种植物的不同个体群都朝着适应各自环境条件的方向发展,导致了不同个体群之间的变异。如果这些变异能够遗传,就形成了在同一个种内分化成不同的个体群类型,即生态型。
同一物种的不同生态型之间不存在生殖隔离,即可以相互杂交。有时所产生的杂交后代比双亲更具活力,这种现象称为杂交优势。农林畜牧业上常利用杂交方式培育新品种。
(七)生态入侵
由于人类有意识或无意识地把某种生物带入适宜于其栖息和繁衍的地区,种群不断扩大,分布区逐步稳定扩展,这种过程称为生态入侵(ecological invasion)。
仙人掌原产美洲,有数百种,其中有26种被引进澳大利亚作为园艺植物。1839年引进作篱笆的Opuntia stricta由于扩展迅速,于1880年被视为“草害”。1890年危害面积已达40000平方千米,1920年为235000平方千米,1925年为243000平方千米。其中有一半面积生长茂盛;高出地面1~2m,行人难以通行。由于防治费用高昂,每公顷约25~100美元,多被废弃为荒田。直到1920年从原产地引进其天敌Cactoblastis cactorum才得以有效控制。
紫茎泽兰(Eupatorium adenophorum)原产墨西哥,1865年作为观赏植物引入夏威夷,1875年引入澳大利亚。后来发展为到处繁衍,泛滥成灾。解放前由缅甸、越南进入我国云南,现已蔓延到25°33′N地区,并向东扩展到广西、贵州境内。它常连接成片,发展为单优势群落,侵入农田,危害牲畜,影响林木生长,成为“害草”。
|
延伸阅读
|